Neurosteroids: Depression, Anxiety and Increasing Production

Neurosteroid disruption has been linked to a wide array of mental and physical health issues, from Alzheimer’s to depression. Neurosteroids are endogenous (originating from within) or exogenous (originating externally) steroids which rapidly modulate neural excitability (how likely a neurone is to fire aka transmit a signal to the next neurone). Although “steroids” are usually associated with images of ‘The Rock’ or ‘The Mountain’ tearing phone books in two and bench pressing small cars while glistening in oil…but I digress, the term steroid really refers to a biologically active organic compound with four rings arranged in a specific molecular configuration. Steroids have two principal biological functions: as essential components of cell membranes which alter membrane fluidity; and as signalling molecules.
What are they?
Neurosteroids are synthesised from cholesterol, which is converted into Pregnenolone and then into all other endogenous steroids. At the inner mitochondrial membrane, cholesterol is converted to Pregnenolone by the action of the P450 cholesterol side-chain cleavage enzyme. Pregnenolone then passes to the fluid inside the cell (cytosol), where it serves as the precursor of all neurosteroids.

In the cytosol, Pregnenolone is then converted, via successive enzymatic steps, to a variety of potent neurosteroids. 5α-reductase type I and 3α-hydroxysteroid dehydrogenase are involved in the biosynthesis of inhibitory (decreasing the likely hood of a neurone firing) neurosteroids, while 3β-hydroxysteroid dehydrogenase and hydroxysteroid sulfotransferases are involved in excitatory (raising the likely hood of a neurone firing) neurosteroid production. (1)
Current research shows that both 5alpha-reductase and 3alpha-HSOR are present in the hippocampus, neocortex and subcortical white matter in both the neurons and glial cells (2). As such, neurosteroids can likely be formed directly in the target brain tissue.
What do they do?
Neurosteroids are potent modulators of the two major neurotransmitter systems that govern Central Nervous System activity — glutamate, the major excitatory neurotransmitter, and gamma-aminobutyric acid (GABA), the major inhibitory neurotransmitter. It appears during states of increased stress that there is a dysregulation of neurosteroids (3) as such potential uses for neurosteroids include mood and anxiety disorders, memory dysfunction, migraines, schizophrenia, alcoholism, sleep disorders, chronic pain, epilepsy, traumatic brain injury and neurodegenerative diseases.

Although the GABA and glutamate neurotransmitter systems and receptors are significant targets for the actions of neurosteroids, these steroids also alter the function of other receptors and ion channels. Alternative targets include nicotinic acetylcholine receptors, glycine receptors, sigma receptors as well as voltage-activated calcium, voltage-activated potassium channels and transient receptor potential. For now, we’re going to focus on GABA and glutamate systems.
GABA

When describing the effects of neurosteroids on GABA-a receptors, it is crucial to have a grasp on some of the complexities of these receptors, especially since not all GABA-a receptor modulators have an antidepressant effect. GABA-a receptors are pentameric chloride channels; these are five (Penta) subunits that come together to form a channel, activated aka gated by the neurotransmitter, GABA. So far there have been 19 GABA-a receptor subunits identified α1–6, β1–3, γ1–3, δ, ε, ρ1–3, π, Ɵ. (Now I know what you’re thinking, why not just name them 1–19? Well now it wouldn’t be science without a bunch of random symbols we no longer use then would it?) Most native GABA-a receptors express three subunits, usually with 2α, 2β and a third subunit, although receptors with only one or two subunit types have been described.
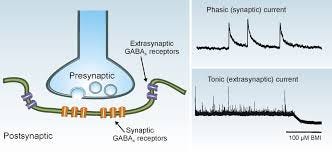
Different types of GABA-a receptors control various forms of inhibition in the brain. GABA-a receptors mediate two distinct forms of inhibition, phasic and tonic.
- The first form consists of fast inhibitory postsynaptic potentials (IPSPs) regulating communication between neurons. IPSPs is a kind of synaptic potential that makes a postsynaptic (one that’s fired) neuron less likely to generate an action potential.
- The second form consists of a persistent inhibitory conductance that plays a crucial role in regulating the membrane potential and network excitability. In the case of phasic inhibition, synaptic GABA-a receptors, facing presynaptic release sites, are activated by brief exposure to a high concentration of GABA. Once released, GABA diffuses throughout the neuropil, the dense feltwork of interwoven cytoplasmic processes of nerve cells, dendrites, axons and of neuroglial cells in the central nervous system and some parts of the peripheral nervous system. It is then taken up by selective plasma membrane transporters, which contribute to the clearance of the neurotransmitter. In the case of tonic inhibition, extrasynaptic GABA-a receptors, localised away from the synapses, are persistently exposed to a low concentration of “ambient” GABA. Phasic (synaptic) inhibition usually involves GABA-a receptors that express γ2 subunits, while more persistent, tonic inhibition is mediated by extrasynaptic receptors that often, although not always, contain a δ-subunit. Receptors mediating tonic inhibition typically differ from those mediating phasic inhibition in their higher sensitivity to GABA, slower ion channel kinetics and weaker desensitisation.
Broadly speaking, neural steroids have one of three effects on GABA-a receptors (GABAaR). Certain neural steroids are GABaAR positive allosteric modulators (PAMs) which increase agonist affinity and/or efficacy, enhancing the actions of GABA in the brain (an agonist is a substance which creates a physiological response when combined with a receptor). The second class of active neural steroids act as negative allosteric modulators (NAMs) which lower agonist affinity and/or efficacy at GABAARs and are activation-dependent, non-competitive inhibitors.
Usually, drugs bind directly to the same part of receptors that the body’s own proteins attach to, this means that those drugs are competing with those endogenous proteins, in effect limiting the potential impact of a drug.
Allosteric modulators get around this by binding to different parts of the receptor. And instead of merely turning on or off the receptor, allosteric modulators can either turn up the volume of the signal being sent by the receptor or decrease the signals. This means that when the body’s naturally occurring protein binds in the receptor, allosteric modulators can either amplify the effect (PAMs) or reduce it (NAMs) depending on which type of allosteric modulators is being administered.
In fact, allopregnanolone is much more effective in potentiating GABA responses compared to benzodiazepines (up to 20-fold compared to 3-fold enhancement at low GABA concentrations) and comparable to barbiturates. (4)
Effects on depression
Now that we’ve examined how these neural steroids act on the brain, we can dive into the more actionable stuff, what can we do actually to help ourselves feel better? We’ll focus on the influence on depression and anxiety.
- A 1998 study by Uzunova, et al. entitled; “Increase in the cerebrospinal fluid content (CSF) of neurosteroids in patients with unipolar major depression who are receiving fluoxetine or fluvoxamine” (a catchy title I know) administered Fluoxetine (0.27–1.5 mg per kg per day for 8 –10 weeks) to 15 patients with unipolar depression, and then checked allopregnanolone levels in their cerebrospinal fluid. They found that treatment with Fluoxetine normalised their allopregnanolone levels and led to a 60% increase from their baseline “The normalisation of CSF allopregnanolone content in depressed patients appears to be sufficient to mediate the anxiolytic (reduction of anxiety) actions of fluoxetine or fluvoxamine via its positive allosteric modulation of GABA type A receptors.” These results have been replicated in a few other studies utilising low doses of Fluoxetine/Prozac.(5)
- A June 2014 study examined daily dosage of Pregnenolone, 500mgs, in patients with Bipolar disorder (often described as depressive episodes along with manic episodes as opposed to unipolar depression). The researchers found “depression remission rates were greater in the pregnenolone group (61%) compared with the placebo group (37%)…Pregnenolone was well tolerated. The results suggest that Pregnenolone may improve depressive symptoms in patients with Bipolar disorder and can be safely administered.” (6)
- Another study involving 70 (37 fully completed the study: 19 pregnenolone, 18 placebo) candidates with bipolar disorder or major depressive disorder were given 100mg/day or placebo for 8 weeks. Participants were assessed using a variety of different scales examining both mood and cognition. Mood was evaluated bi-weekly, while cognition was assessed at baseline, and weeks 4 and 8. “Change per week in the pregnenolone-treated patients was over twice that of the placebo. Those who [completed the study] showed a significantly greater decrease in depressive symptoms with Pregnenolone.” Cognition was unaffected. (7)
Effects on Anxiety
Along with modulating depressive symptoms, neurosteroids appear to have a positive impact on anxiety which makes sense after examining their close relationship to GABA receptors.
- A 2013 study of; 31 people, 16 active and 15 placebo, were given 400mgs of Pregnenolone, administered orally, and underwent a 3T fMRI while performing the Shifted-Attention Emotion Appraisal Task (which probes emotional processing and regulation). They concluded that “Pregnenolone administration reduced activity in neural circuits associated with the generation of negative emotions [decreasing right activity in both the amygdala and insula]”, “increased activity in the dmPFC, a region linked to regulatory control over emotion.”, “increased connectivity between amygdala and dmPFC during the appraisal, with greater connectivity associated with reduced self-reported anxiety.” (8)
- Further, a study from 1994 investigated the relationship between Corticotropin-releasing hormone (CRH) and allopregnanolone. CRH plays a significant role in stress-induced activation of the HPA axis. The hypothalamic–pituitary–adrenal axis (HPA axis or HTPA axis) is comprised of the hypothalamus, the pituitary gland and the adrenal glands. Together these have a significant influence on how the nervous system and endocrine system interact in response to stress. During stress, CRH levels are increased in the hypothalamus, resulting in increased release of cortisol and dehydroepiandrosterone (DHEA). Sustained hyperactivity corticotropin-releasing hormone-producing neurons are thought to be related to both anxiety and affective disorders. Researchers observed, from examining both hypothalamic organ explants in-vitro (meaning “test-tube experiments”) and gene expression of CRH in the hypothalamic paraventricular nucleus (a cluster of neurones in the hypothalamus) of rats that “[Allopregnanolone] has anxiolytic effects that are mediated through interactions with hypothalamic corticotropin-releasing hormone in both, genomic and non-genomic fashions” (9)
- A small trial comparing 8 men with anxiety disorders and 8 healthy people proposed a link between anxiety and lower levels of pregnenolone sulfate: “Although the lower level of allopregnanolone in patients was only statistically suggestive, the direction of the results would indicate a decrease in an endogenous anxiolytic agent and lower agonist activity at the level of the GABA/BZ receptor in patients with anxiety disorders.” (10)
Increasing neurosteroid levels
Now the important part is, how do we utilise this information in a meaningful way? What can we trial to boost our overall levels of these potent anxiolytic compounds?
Pregnenolone Supplementation
To me, the obvious place to start is via direct supplementation of Pregnenolone. Returning to the 2009 “Proof-of-Concept Trial with the Neurosteroid Pregnenolone Targeting Cognitive and Negative Symptoms in Schizophrenia” researchers noted that: “Treatment with Pregnenolone resulted in fourfold elevations in serum levels of Pregnenolone…tripled serum pregnenolone sulfate levels…and increased the GABAergic neurosteroid allopregnanolone fivefold. Pregnenolone administration also increased serum progesterone over fourfold and DHEAS levels by approximately 16% (Table 6). Treatment with Pregnenolone did not increase serum testosterone, free testosterone, cortisol, DHEA, estradiol, or androstenedione levels.” (11) Bearing in mind the small duration (8 weeks) and the small sample size (n=18) the findings demonstrate that, up to 400mgs per day of orally supplemented Pregnenolone are safe and present a marked increase in serum levels of Pregnenolone. But more importantly, a 500% increase in allopregnanolone and didn’t alter other hormones.

Another study, “A Randomised, Double-Blind, Placebo-Controlled Trial of Pregnenolone for Bipolar Depression” with a larger sample size (n=80), this time in depressed patients with a bipolar diagnosis, conferred a similar result. With pregnenolone supplementation (500mgs) not only being tolerated equally well as the placebo but resulted in “remission rates were greater in the pregnenolone group (61%) compared with the placebo group (37%).” (6)
Palmitoylethanolamide (PEA)

Palmitoylethanolamide (PEA) is an endocannabinoid-like compound and an endogenous fatty acid amide playing a role in intracellular signalling. Part of PEA’s pharmacological effects is mediated through the ligand-activated nuclear hormone family peroxisome proliferator-activated receptors (PPARs). PPAR agonism is involved in a variety of pathways, including neurological inflammation and oxidative stress. Studies in astrocytes and the spinal cord showed that PEA binding to PPAR-stimulated the biosynthesis of allopregnanolone, through increased expression of the steroidogenic acute regulatory protein (StAR) and cytochrome P450 side-chain cleavage (12). One study conducted, injected mice with PEA with between 5–20mgs per kg, this resulted in an increase of allopregnanolone throughout the brain;
The olfactory bulb saw an increase of: 5 mg/kg resulted in an +164%, 10 mg/kg +185%, 20 mg/kg: +160%
The hippocampus had an increase of 138% with a 5mg/kg dose, and 178% with a 10 and 20mg dose
The amygdala had increased allopregnanolone levels of 34% at 5mg/kg, 42% at 10mg/kg, and 120% at 20mg/kg
A 2011 in-vitro study looked at PEA’s effects on mouse astrocytes. These are cells in the brain and spinal cord involved in a litany of biological functions. In this study, they concluded that there was “an increase in steroidogenic acute regulatory protein (StAR) and cytochrome P450 enzyme (P450scc) expression, both comprising proteins considered to be involved in crucial steps of neurosteroid formation.” Further, the researchers noted that allopregnanolone levels were increased in the PEA-treated astrocytes. PEA also showed a protective effect via a reduction in the malondialdehyde build up. Malondialdehyde is frequently used as a biomarker of oxidative stress. PEA was partially inhibited by finasteride, a drug which inhibits the enzyme 5α-reductase, which is involved in the transformations of a variety of endogenous steroids including allopregnanolone, suggesting that the reduction of oxidative stress is mediated through allopregnanolone synthesis. (13)
Other avenues
Above are the main two compounds I could find related to increased neurosteroid production, which had, not only rodent models but, some human studies with supporting evidence. Everything else I could find was limited to cell lines and purely rodent data. Most of these examined pathways relevant to neurosteroid production; however, their production not being the endpoint. Most of these referenced steroidogenic acute regulatory protein (StAR), critical to the entry of cholesterol into the mitochondria. Once inside the mitochondria, cholesterol is metabolised by the steroidogenic enzyme Cyp11a1 to Pregnenolone, which is further converted to various other steroids we’ve discussed.
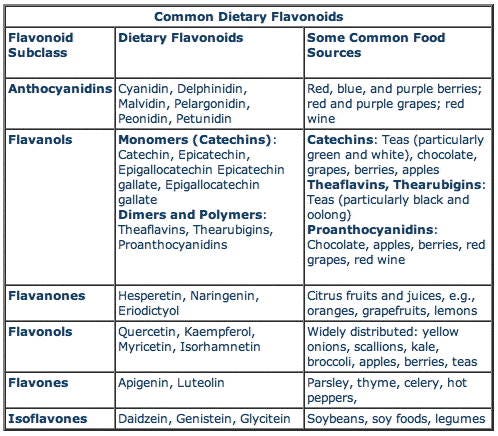
A variety of flavonoids were the main compounds that came up during my searches. Flavonoids are widely distributed in plants, fulfilling many functions; UV filtration, symbiotic nitrogen fixation and floral pigmentation etc. They are found in high densities in many plant products including; parsley, onions, blueberries and other berries, black tea, green tea and oolong tea, bananas, citrus fruits, red wine, sea-buckthorns, buckwheat, and dark chocolate (a cocoa content of ≥70%).
- “Flavonoids, such as chrysin, apigenin, luteolin, and quercetin, have been shown to enhance StAR gene expression and steroidogenesis in Leydig cells by blocking the cyclooxygenase-2 (Cox2)-dependent signalling. In ageing males, Cox2 [which is involved in inflammation via conversion of arachidonic acid to prostaglandin H2] is increased in Leydig cells, resulting in depressed Star gene expression and testosterone production” (14)
- “The steroidogenic effect of quercetin was studied with MA-10 mouse Leydig cells. It was reported that quercetin increased StAR mRNA levels, StAR promoter activity and steroid hormone production…Following these studies, the transcription factors in StAR gene transcription were examined. It was found that the incubation of MA-10 Leydig cells with quercetin reduced DAX-1 protein, similar to the effects of other flavonoids. The reduction in DAX-1 protein was associated with a dramatic increase in StAR protein (Fig. 3C), suggesting that quercetin enhance StAR gene expression by reduction of DAX-1 expression.” (15)
So flavones had a stimulatory effect on testicular Leydig cell function by increasing steroidogenic gene expression via limiting Cox2 and DAX-1 expression.
- “These results show that the GABA-binding site at GABAA receptors participates in the acute antidepressant-like effects of chrysin, similar to neurosteroids, in ovariectomised rats.” (16)
Again the flavonoid, chrysin, had a similar effect on rodents to progesterone and allopregnanolone.
Conclusions and summary
Hopefully, I’ve done at least a mediocre job of explaining how neurosteroid production is altered in many mood-disorders and offers a unique and novel therapeutic avenue. Although most exogenous pharmaceutical modulators are in the early phases of human trials, there does seem to be some natural alternatives that can be implemented now. Alternatives which not only may help with neural steroid production and regulation but that also have other beneficial health impacts. Personally, how I use this information will be to;
1. Palmitoylethanolamide x 1.5g per day
2. Pregnenolone x 500mgs per day
3. Aiming for a wider range of fruits and vegetables and consume more, kale, spinach, berries, grapefruit, cacao and celery
4. And maybe rotate through some different flavonoid supplements
Although we have emphasised the importance of GABA (and to a point glutamate) receptors, neurosteroids have other synaptic and extrasynaptic “targets” that could contribute to their psychotherapeutic actions, including potent effects on different receptors and channels. So there’s plenty more information out there, and I’d advise if you’re interested in reviewing some of the papers referenced below.
Glossary
Find more on
Personal website =
